Жизнедеятельность и значение уксуснокислых бактерий
Содержание:
- Гомоферментативное молочнокислое брожение
- Немного истории
- Полезные свойства
- Уксуснокислые бактерии
- Использование человеком
- Среда обитания
- Лимоннокислое брожение
- 1.2.5 Дрожжи
- 11.6 Окисление жиров и высших жирных кислот микроорганизмами. Микроорганизмы — возбудители порчи жиров
- Биохимия
- Негативные свойства
- Маслянокислое брожение
- Строение бактерий
- Уксуснокислые бактерии
Гомоферментативное молочнокислое брожение
Гомоферментативное молочнокислое брожение представляет собой энергетическую сторону образа жизни группы гомофермен-тативных молочнокислых бактерий. Черты древности этой группы видны не только в процессе добывания ее представителями энергии, но и в других сторонах их метаболизма, о чем будет сказано в разделе, посвященном краткой характеристике этих бактерий.
Гомоферментативное молочнокислое брожение, в основе которого лежит гликолитический путь разложения глюкозы, является единственным способом получения энергии для группы эубак-терий, которые при сбраживании углеводов превращают в молочную кислоту от 85 до 90 % сахара среды. Бактерии, входящие в данную группу, морфологически различны. Это кокки, относящиеся к родам Streptococcus и Pediococcus, а также длинные или короткие палочки из рода Lactobacillus. Последний подразделяется на три подрода.
|
Схема регенерации окисленного НАД в аэробных ( А и анаэробных условиях. В — молочнокислое брожение. С — спиртовое брожение.| Виды брожений, основанные на гликолизе. |
Гомоферментативное молочнокислое брожение идентично по химизму реакциям гликолиза в анаэробных условиях.
Гомоферментативное молочнокислое брожение вызывают бактерии рода Lactobacillus и стрептококки. Они могут сбраживать различные сахара с 6 — ю ( гексозы) или 5 — ю ( пентозы) углеродными атомами, некоторые кислоты. Однако круг сбраживаемых ими продуктов ограничен.
В процессе гомоферментативного молочнокислого брожения синтезируются 2 молекулы АТФ на 1 молекулу сброженной глюкозы; в процессе дыхания при полном окислении молекулы глюкозы образуется 38 молекул АТФ. В обоих случаях эффективность запасания выделяющейся энергии в макроэргических связях АТФ приблизительно одинакова.
Возникнув как первый, далекий от совершенства энергетический процесс, гомоферментативное молочнокислое брожение не было потом отброшено в процессе эволюции. Наоборот, оно закрепилось и существует сейчас в виде гликолиза у подавляющего большинства прокариот, дрожжей, грибов, а также у высших животных и растений, но только как первый этап более совершенного энергетического процесса, сформировавшегося в результате последующего развития способов получения энергии живыми организмами. Чем объясняется такая судьба гомоферментативного молочнокислого брожения. Вероятно, оказалось выгодным использовать его в качестве первого подготовительного этапа по следующим причинам: 1) высокая энергетическая эффективность ( не путать с энергетическим выходом процесса.
Собственно гликолиз — это определенная последовательность ферментативных реакций от углевода до пировиноградной кислоты, поэтому, строго говоря, гликолиз не является синонимом гомоферментативного молочнокислого брожения, но 10 из 11 реакций у этих процессов идентичны.
|
Схема энергетических и транспортных процессов у молочнокислых бактерий. Темный кружок — переносчик. В — молекула растворенного вещества. глюкоза поступает в клетку с помощью фосфотрансфе-разной системы. Остальные объяснения в тексте. |
Обратимо функционирующие протонные АТФазы мы находим у первичных анаэробов, получающих энергию в процессе брожения. Обнаружено, что выделение во внешнюю среду молочной и уксусной кислот молочнокислыми бактериями и клостридиями приводит к созданию на ЦПМ протонного градиента. У стрептококков, осуществляющих гомоферментативное молочнокислое брожение, молочная кислота накапливается в клетке в виде аниона, для которого ЦПМ практически непроницаема.
Возникнув как первый, далекий от совершенства энергетический процесс, гомоферментативное молочнокислое брожение не было потом отброшено в процессе эволюции. Наоборот, оно закрепилось и существует сейчас в виде гликолиза у подавляющего большинства прокариот, дрожжей, грибов, а также у высших животных и растений, но только как первый этап более совершенного энергетического процесса, сформировавшегося в результате последующего развития способов получения энергии живыми организмами. Чем объясняется такая судьба гомоферментативного молочнокислого брожения. Вероятно, оказалось выгодным использовать его в качестве первого подготовительного этапа по следующим причинам: 1) высокая энергетическая эффективность ( не путать с энергетическим выходом процесса.
Окислительно-восстановительные превращения имеют место на двух этапах процесса, именно они приводят к получению клеткой энергии. Это результат того, что процесс замкнут на себя, т.е. субстрат является и источником веществ — доноров электронов и источником веществ — их акцепторов. Все это, вместе взятое, определило судьбу гомоферментативного молочнокислого брожения.
Немного истории
Согласно археологическим свидетельствам, бактериальные микроорганизмы были на земле еще 3,5 миллиарда лет назад. Эукариоты же (клетки, содержащие ядро), появились около 1,4 миллиарда лет назад.
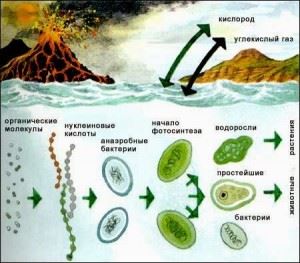
Жизнедеятельность бактерий возможна при большой вариации природных условий. Это горячие источники, где температура воды более 90 градусов, а также среды с температурой ниже ноля. Они являются единственными жителями соленого Мертво моря. Запыленный воздух городов содержит их больше, чем свежий сельский. Их гораздо меньше в воздухе холодных полюсов, однако они есть. Обнаружены бактериальные включения даже на высоте 8 км от Земли.
Выявление бактерий в пищеварительной системе здоровых животных, людей показало их большое значение для нормального ее функционирования. Для жвачных животных они жизненно необходимы для процесса переваривания растительной пищи.
Полезные свойства
Помимо того, что жизнедеятельность бактерий наносит вред виноделию, можно выделить и целый ряд примеров удачного использования человеком характеристик микроорганизмов.
Так, главная роль уксуснокислым бактериям отведена в производстве и изготовлении столового уксуса из вина или разбавленного спирта. Осуществляется это по сей день двумя способами.
Первый представляет собой более медленный, но тщательный процесс, называемый орлеанским или просто французским. Для него необходимо подготовить вино, предварительно подкисленное или разбавленное водой. Поместить его в подготовленные плоские емкости, чтобы поверхность соприкосновения с воздухом была максимальной, и выпустить в жидкость частицы уже образованной ранее пленки Acetobacter orleanense. Она имеет желтый цвет и прочную текстуру, позволяющую сохранить прозрачность жидкости под ней.
После окончания брожения из емкости аккуратно забирают часть субстрата и заменяют его аналогичным количеством разбавленного вина, после чего процесс возобновляется.
Второй способ более быстрый и применим для окисления разбавленного спирта. Для этого его пропускают через специальные емкости с буковыми стружками, чтобы также увеличить поверхность сцепления с бактериями. Емкости при этом обязательно снабжены ложными доньями с возможностью пропускать через них воздух. Таким образом, разбрызгиваемый потоками воздуха спирт оседает на стружке и окисляется, после чего его отбирают из сосуда снизу, а сверху доливают новый субстрат.
Кроме этого клетки используются при:
- мочении яблок вместе с дрожжами;
- производстве аскорбиновой кислоты;
- выращивании чайного гриба;
- изготовлении кефира.
Вообще, в производстве абсолютно всех молочнокислых продуктов наблюдается параллельное брожение, то есть молочнокислые и уксуснокислые бактерии вместе обеспечивают появление продуктов переработки молока в таком виде, как мы и привыкли.
Уксуснокислые бактерии
Являются вредной микрофлорой, вызывающей уксусное скисание вина. Уксуснокислые бактерии принадлежат к роду Acetobacter. Они имеют палочковидную форму. Клетки короткие, толстые, заключены в капсулу, располагаются в жидкой среде попарно, редко одиночно, иногда в виде цепочек.
Некоторые уксуснокислые бактерии подвижны. Характеризуются высокой скоростью размножения: при благоприятных условиях из одной клетки за 12 часов может образоваться 17 млн. бактерий.
Свое название уксуснокислые бактерии получили из-за способности окислять этиловый спирт в уксусную кислоту при свободном доступе кислорода воздуха.
Уксуснокислые бактерии легко и быстро размножаются на поврежденных ягодах винограда; попав в сусло, при брожении его они не погибают. Для своего роста и развития они нуждаются в питательных веществах: углероде, азоте (в основном усваивают его из аминокислот), витаминах. Все уксуснокислые бактерии хорошо используют в качестве источника углерода моносахариды, многоатомные спирты, могут усваивать кислоты, в том числе вырабатываемую ими уксусную кислоту.
Такое явление называется переокислением. Энергию уксуснокислые бактерии получают за счет реакций окисления. Окисление бактериями этилового спирта в уксусную кислоту сопровождается образованием этилацетата, который придает винам неприятные тона во вкусе и аромате, характерные для уксуснокислого скисания. Из 1 % об. этанола образуется 1 г уксусной кислоты.
Помимо этилового спирта, уксуснокислые бактерии окисляют другие одноатомные спирты, а именно: пропиловый спирт — в пропионовую кислоту, бутиловый — в масляную, изоамиловый — в изовалериановую кислоту, а также многоатомные спирты— сорбит, глицерин, маннит.
При развитии уксуснокислых бактерий на поверхности вина, виноградного сока, других жидких продуктов переработки винограда образуются слизистая пленка или пристенное кольцо. Через некоторое время возможно погружение пленки в жидкость. Характерной особенностью пленки из уксуснокислых бактерий является ее способность всползать на стенки стеклянной посуды. Чаще всего бактерии образуют пленку совместно с пленчатыми дрожжами родов Candida, Pichia, Hansenula.
На развитие уксуснокислых бактерий большое влияние оказывает температура. Для них благоприятен широкий диапазон: от 10 до 35 °С. Бактерии сохраняются при более низких температурах, но погибают при более высоких в зависимости от величины рН, концентрации сернистой кислоты и других факторов. Так, в столовом вине при отсутствии кислорода клетки вида Acetobacter aceti погибают в течение 10 минут при температуре 50 °С.
С повышением крепости столовых вин активность уксуснокислых бактерий снижается, однако при температуре 20—25°С бактерии способны развиваться и подвергать скисанию вина крепостью 14—16% об.
Уксуснокислые бактерии чувствительны к SO2: при содержании его в количестве 125—150 мг/л жизнедеятельность бактерий приостанавливается при температуре 15—18 °С только на 10 дней; при введении в виноматериал S02 в количестве 50 мг/л в анаэробных условиях они теряют свою жизнедеятельность при температуре 10°С и ниже на 5—10-е сут, а при 28— 35 °С — на несколько часов. Для инактивации всех видов уксуснокислых бактерий необходимо сульфитировать вина до содержания в них общего количества S02 не менее 175 мг/л.
Уксуснокислые бактерии развиваются в вине, соках, слабоалкогольных напитках, в не полностью долитых или недостаточно плотно закрытых емкостях, при пористой клепке бочек, при брожении мезги, в таре с остатками вина. Вино, в котором брожение закончилось, надо хранить без доступа воздуха.
При хранении вин в металлических и железобетонных емкостях, заполненных ниже установленных норм (недолитых), рекомендуется использовать герметизирующий состав СВС с 2% метабисульфита калия. Производство виноградных соков и напитков на их основе базируется на использовании пастеризации — кратковременного нагрева продукта в бескислородных условиях при температуре 55—75 °С и выше. В целях профилактики рекомендуется периодически производить дезинфекцию помещений, тары и коммуникаций.
Использование человеком
Основная польза от брожения — это превращение, например, сока в вино, зерна и других исходных продуктов в пиво, а углеводов в диоксид углерода при приготовлении хлебного теста. Широко используется человеком также молочнокислое брожение для приготовления кисломолочных продуктов, квашения овощей и приготовления силоса.
Поскольку фрукты сбраживаются в своем натуральном состоянии, брожение как процесс изменения пищевых продуктов появилось раньше человеческой истории. Однако люди с некоторых пор научились контролировать процесс брожения. Есть веские доказательства того, что люди сбраживали напитки в Вавилоне около 5000 г. до н. э., в Древнем Египте около 3000 г. до н. э., в доиспанской Мексике около 2000 г. до н. э. и в Судане около 1500 г. до н. э. Также существуют данные о дрожжевом хлебе в Древнем Египте около 1500 г. до н. э. и сбраживании молока в Вавилоне около 3000 г. до н. э. Китайцы, вероятно, первыми стали сбраживать овощи.
По Штейнкраузу (Steinkraus; 1995), брожение пищи выполняет пять главных задач:
- Обогащение видов пищи разнообразием вкусов, ароматов и текстуры
- Сохранение существенного количества пищи с помощью молочной кислоты, алкоголя, уксусной кислоты и щелочного брожения
- Биологическое обогащение пищи протеинами, важными аминокислотами, важными жирными кислотами и витаминами
- Детоксификация в процессе брожения пищи
- Уменьшение времени и затрат на приготовление пищи
У брожения есть несколько преимуществ, важных для приготовления или сохранения пищи. В процессе брожения можно получать важные питательные вещества или устранять непитательные. С помощью брожения пищу можно дольше сохранять, поскольку брожение может создать условия, неподходящие для нежелательных микроорганизмов. Например, при квашении кислота, получаемая из доминирующей бактерии, препятствует росту всех других микроорганизмов.
Среда обитания
Уксуснокислые бактерии, как показывает характеристика их питания, являются обитателями сред брожения. Чаще всего они размножаются сразу же за дрожжами, используя выделяемый ими спирт. Поэтому в природе их можно обнаружить на следующих продуктах:
- сахаросодержащие спелые и перезрелые фрукты и овощи;
- зрелый виноград, особенно загнившие гроздья, что отлично видно на фото, сделанных даже без помощи увеличения;
- цветочный нектар и другие.

Особенно много их в скисших фруктовых соках, непастеризованном пиве, вине, спиртовой бражке, сидре и т.д. Как уксуснокислые, так и молочнокислые бактерии присутствуют в кефире и других кисломолочных продуктах, являются неотъемлемыми участниками процесса квашения и засолки овощей, мочения яблок.
Большое значение для их развития имеет температура. Практически для всех видов этого семейства бактерий нижним пределом является –6…–10°С. Верхняя граница составляет +35…+45°С. Температурные пределы непостоянны, так как они находятся во взаимосвязи с прочими характеристиками среды. Поэтому в природе бактерии отлично себя чувствуют на богатых питательными веществами продуктах, в которых происходит брожение, при постоянном доступе свободного кислорода.
Принято считать, что определяющее значение в распространении этих бактерий имеет плодовая мушка дрозофила, как можно увидеть на фото, и уксусные угрицы (вид круглых червей).

Лимоннокислое брожение
Лимоннокислое
брожение – это
процесс окисления сахаров до лимонной
кислоты в результате жизнедеятельности
плесневых грибов.
В 1891 г. немецкий
ученый Вемер установил способность
плесневых грибов продуцировать
органические кислоты. А благодаря
работам Бутковича, Костычева, а также
их учеников в нашей стране в начале 30-х
гг. было создано микробиологическое
производство лимонной кислоты с помощью
плесневого гриба Aspergillusniger.
Последние два десятилетия в качестве
продуцентов все шире применяются
экспериментально полученные мутантные
штаммы плесневых грибов, отличающиеся
от природных рядом положительных
свойств, в первую очередь более высоким
выходом целевого продукта.
1.2.5 Дрожжи
Дрожжи
– это высшие грибы, утратившие способность
образовывать мицелий и превратившиеся
в одноклеточные организмы
Относятся
к надцарству эукариот, отделу истинных
грибов, большинство дрожжей являются
представителями двух классов: аскомицетов
и дейтеромицетов. Кроме того, дрожжи
делятся на спорогенные и аспорогенные.
В молоке и молочных продуктах чаще всего
встречаются спорогенные дрожжи семейства
Saccharomycetaceae (например, родов Saccharomyces,
Zygosaccharomyces) и аспорогенные дрожжи семейства
Torulopsidaceae
(родов Torulopsis,
Candida,
Mycoderma
и др.).
В
основу классификации дрожжей положены
следующие признаки: различия в характере
их вегетативного размножения, способность
к спорообразованию и половому размножению,
а также другие морфологические и
физиологические свойства.
Многие
дрожжи являются возбудителями спиртового
брожения – процесса анаэробного
окисления сахаров до этилового спирта.
Возможность
дрожжей размножаться в молоке и молочных
продуктах определяется их способностью
сбраживать или окислять лактозу, а также
наличием в молоке микрофлоры, обладающей
(-галактозидазной активностью. В связи
с этим дрожжи, встречающиеся в молоке
и молочных продуктах, делятся на 3 группы:
1.
Дрожжи, не способные к спиртовому
брожению, но потребляющие лактозу путем
непосредственного окисления (в молоке
растут, но лактозу не сбраживают). К
таким дрожжам относятся дрожжи родов
Mycoderma, Torula.
2.
Дрожжи не сбраживающие лактозу, но
сбраживающие другие сахара. Эти дрожжи
могут развиваться только в совместной
культуре с микроорганизмами, которые
обладают (-галак-тозидазной активностью
и осуществляют гидролиз молочного
сахара до глюкозы и галактозы. Такими
дрожжами являются большинство видов
дрожжей рода Saccharomyces.
3.
Дрожжи, сбраживающие лактозу. Таких
дрожжей не много. Наиболее часто в
молочных продуктах встречаются следующие
виды дрожжей этой группы: Saccharomyces
lactis,
Saccharomyces
fragilis,
Torulopsis
kefir,
Torulopsis
sphaerica,
Candida
pseudotropicalis
и др.
Большинство
дрожжей являются факультативными
анаэробами, некоторые дрожжи – аэробы.
Хорошо растут в кислой среде (ацидофилы).
По отношению к температуре дрожжи
являются мезофилами, так как оптимальная
температура для их развития 25-300С. Более
высокая температура стимулирует развитие
дрожжей вида Torulopsis sphaerica и дрожжей, не
сбраживающих лактозу. В качестве
источника углерода лучше всего используют
гексозы, другие углеводы, спирты,
органические кислоты. Источниками азота
для них являются соли аммония, аминокислоты,
пептиды.
Естественным
местообитанием дрожжей является
поверхность плодов и ягод, сок и
поверхность листьев, нектар, вода, почва,
кожные покровы и пищеварительный тракт
людей и животных. Имеются патогенные и
условно-патогенные формы дрожжей,
которые вызывают кандидомикозы.
Роль
дрожжей в формировании качества молочных
продуктов исключительно велика. Они
используются при производстве кефира
и кумыса, являясь не только возбудителями
спиртового брожения, но и продуцентами
витаминов группы В, антибиотических
веществ, подавляющих развитие туберкулезной
палочки и других патогенных микроорганизмов.
Продукты жизнедеятельности дрожжей
активизируют развитие молочнокислых
бактерий. Некоторых дрожжи используются
в производстве масла, так как предотвращают
развитие на его поверхности микроскопических
грибов и, таким образом, повышают
стойкость масла в процессе хранения.
С
другой стороны, дрожжи являются
вредителями производства многих молочных
продуктов. Интенсивное развитие дрожжей
незаквасочного происхождения нередко
приводит к вспучиванию, изменению вкуса
творога, сметаны, сладких творожных
изделий, обильному газообразованию
сгущенного молока с сахаром (бомбаж
банок), возникновению спиртового вкуса
и запаха, а также к вспучиванию сыров9.
11.6 Окисление жиров и высших жирных кислот микроорганизмами. Микроорганизмы — возбудители порчи жиров
Жиры представляют
собой сложные эфиры глицерина и высших
жирных кислот.
Так
как жиры – высокомолекулярные соединения,
то в неизменном виде внутрь клетки они
попасть не могут. Поэтому вначале
происходит гидролиз жира при участии
фермента липазы, которая имеется у
многих микроорганизмов. В результате
образуются глицерин и высшие жирные
кислоты. Этот процесс не обеспечивает
клетки энергией, поэтому образовавшиеся
продукты гидролиза используются
различными микроорганизмами в качестве
энергетического материала. Процесс
протекает только в аэробных условиях.
Глицерин подвергается
окислению уксуснокислыми бактериями
до диоксиацетона и далее микроскопическими
грибами до углекислого газа и воды.
Высшие
жирные кислоты окисляются труднее и
медленнее. В процессе окисления образуются
промежуточные продукты: кетоны, альдегиды,
оксикислоты и др., которые придают
окисленному жиру прогорклый вкус.
Возбудители.
Наиболее активными микроорганизмами
в процессе разложения жира являются
бактерии рода Pseudomonas, особенно
флуорисцирующие (продуцирующие пигменты)
и мицелиальные грибы: Oidium lactis, многие
виды Aspergillus, Penicillium.
Практическое
значение процесса
Процесс
разложения жиров отмерших животных и
растенийпроисходитпостоянно
и имеетбольшое
значение вкруговороте
веществ в природе.
С
другой стороны, в пищевой промышленности
микроорганизмы, окисляющие жиры, приносят
вред, вызывая порчу пищевых жиров и
жира, содержащихся в различных пищевых
продуктах.
Следует
учитывать, что многие жирорасщепляющие
микроорганизмы являются психрофилами,
поэтому способны развиваться при
хранении пищевых продуктов в охлажденном
состоянии.
Биохимия
Брожение — это процесс, важный в анаэробных условиях, в отсутствие окислительного фосфорилирования. В ходе брожения, как и в ходе гликолиза, образуется АТФ. Во время брожения пируват преобразуется в различные вещества.
Хотя на последнем этапе брожения (превращения пирувата в конечные продукты брожения) не освобождается энергия, он крайне важен для анаэробной клетки, поскольку на этом этапе регенерируется никотинамидадениндинуклеотид (NAD+), который требуется для гликолиза
Это важно для нормальной жизнедеятельности клетки, поскольку гликолиз для многих организмов — единственный источник АТФ в анаэробных условиях.. В ходе брожения происходит частичное окисление субстратов, при котором водород переносится на NAD+ (никотинамидадениндинуклеотид)
В ходе других этапов брожения его промежуточные продукты служат акцепторами водорода, входящего в состав NADH; в ходе регенерации NAD+ они восстанавливаются, а продукты восстановления выводятся из клетки.
В ходе брожения происходит частичное окисление субстратов, при котором водород переносится на NAD+ (никотинамидадениндинуклеотид). В ходе других этапов брожения его промежуточные продукты служат акцепторами водорода, входящего в состав NADH; в ходе регенерации NAD+ они восстанавливаются, а продукты восстановления выводятся из клетки.
Конечные продукты брожения содержат химическую энергию (они не полностью окислены), но считаются отходами, поскольку не могут быть подвергнуты дальнейшему метаболизму в отсутствие кислорода (или других высокоокисленных акцепторов электронов) и часто выводятся из клетки. Следствием этого является тот факт, что получение АТФ брожением менее эффективно, чем путём окислительного фосфорилирования, когда пируват полностью окисляется до диоксида углерода. В ходе разных типов брожения на одну молекулу глюкозы получается от двух до четырёх молекул АТФ (ср. около 36 молекул путём аэробного дыхания).
Негативные свойства
Несмотря на такое положительное значение, уксуснокислые бактерии предполагают и опасность для определенных отраслей. Так, в виноделии микроорганизмы считаются патогенными, поскольку способны провоцировать процесс брожения, несмотря на то что они практически всегда содержатся в пиве и вине. Для их деактивации следует тщательно укупоривать напитки от контакта с воздухом, ведь именно он и провоцирует начало брожения. Полученная от закисания вина уксусная кислота всегда собирается на дне, откуда ее можно собрать, но вкус и аромат оставшегося напитка уже навсегда будет испорчен.

Если начавшийся процесс игнорировать, то все содержимое емкости может превратиться в обычный винный уксус.
Аналогичным образом бактерии способны вызывать закисание засоленных или маринованных овощей.
Маслянокислое брожение
Маслянокислое брожение осуществляется в большинстве случаев облигатными анаэробами, т. е. организмами, способными существовать только в бескислородной среде.
В ходе маслянокислого Б. образуются не только масляная к-та, но в некоторых случаях и весьма значительные количества этилового спирта, молочной н уксусной кислот, а также газообразного водорода и углекислого газа. С помощью маслянокислого Б. осуществляется разложение органических веществ в условиях недостатка или полного отсутствия кислорода (болота, заболоченные места). Большое промышленное значение имеет маслянокислое Б. пектиновых веществ, происходящее при замочке стеблей льна, конопли и получении волокон. Вместе с тем деятельность бактерий, осуществляющих этот вид Б., необходимо предотвращать при приготовлении различного рода пищевых продуктов во избежание ухудшения вкуса и порчи последних (напр., прогоркание сливочного масла, силоса и т. п.).
Спиртовое, молочно- и маслянокислое Б.— основные типы Б.; остальные многочисленные виды Б. представляют собой либо различные их сочетания, либо осуществляются на базе тех или иных продуктов, возникающих в ходе основного вида Б. Так, в результате уксуснокислого брожения происходит окисление этилового спирта при участии кислорода воздуха. Этот вид Б. осуществляется специфическими уксуснокислыми бактериями. Суммарное уравнение уксуснокислого Б.:
CH3CH2OH + O2 = CH3COOH + H2O.
По исчерпании запасов спирта бактерии окисляют образованную им уксусную к-ту до углекислого газа и воды.
К Б., осуществляющемуся с участием О2, относится глюконовокислое брожение — образование глюконовой к-ты из глюкозы:
C6H12O6 + H2O + O2 → CH2OH(CHOH)4COOH + H2O2.
Оно вызываемся нек-рыми бактериями и плесневыми грибами. Глюконовая к-та — ценное соединение, широко применяемое в медицине и фарм, промышленности (см. Глюконовая кислота).
Лимоннокислоe брожениe осуществляется нек-рыми представителями плесневых грибков; особенно эффективны отдельные штаммы Aspergillus niger. Исходным продуктом служит Пировиноградная к-та, превращение к-рой идет одновременно в двух направлениях. Часть ее окисляется в уксусную, тогда как другая, присоединяя углекислоту, образует щавелевоуксусную к-ту. При конденсации уксусной и щавелевоуксусной кислот образуется лимонная к-та. Помимо лимонной к-ты, при лимоннокислом Б. образуются бутиловый спирт, ацетон, а также этиловый спирт, углекислый газ и водород.
Бутанолово-ацетоновое брожение осуществляют анаэробные бактерии Clostridium acetobutylicum. Главные продукты, образующиеся в ходе этого вида Б.,— н-бутиловый спирт, ацетон, этиловый спирт, углекислота, водород. Ацетоуксусная к-та (CH3COCH2COOH) и образующийся при ее декарбоксилировании ацетон (CH3COCH3), а также β-оксимасляная к-та составляют группу так наз. ацетоновых тел (см. Кетоновые тела), которые накапливаются в крови и моче животных при различных патологических состояниях и заболеваниях (диабет, голодание). В нормальных же условиях эти соединения окисляются с образованием безвредных для организма углекислоты и воды.
Высокая экономическая эффективность, чистота получаемых при Б. ценных продуктов лежат в основе все более широкого использования Б. в самых различных отраслях народного хозяйства.
Библиография: Кретович В.Л. Основы биохимии растений, М., 1971; Малер Г. иКордес Ю. Основы биологической химии, пер. с англ., М., 1970; Рубин Б. А. Курс физиологии растений, М., 1971;Рэкер Э. Биоэнергетические механизмы, пер. с англ., М., 1967. библиогр.; Шапошников В. Н. Техническая микробиология, М., 1948; H a s s i d W. Z. Transformation of sugars in plants, Ann. Rev. plant Physiol., v. 18, p. 253, 1967, bibliogr.
Строение бактерий
Размеры этих микроорганизмов настолько малы, что стандартным световым микроскопом с трудом различимы мельчайшие из них.
В своем строении бактерии обладают только одной ДНК, которая крепится своим концом к стенке клетки. Остальные органеллы также не имеют оболочек. В выполнении физиологической митохондриальной функции по синтезу энергии значение имеет вся клетка.
Бактериальная клетка снаружи имеет стенку, сходную по строению с таковой у клеток растений, однако состоит она из других полимеров. Стенку покрывает капсула. Некоторые бактерии снабжены жгутиками для передвижения.
Уксуснокислые бактерии
Уксуснокислые бактерии, выделенные в роды Gluconobacter и Acetobacter, могут получать энергию, осуществляя неполное окисление ряда органических соединений. Это грамотрицательные бесспоровые палочки, слабоподвижные за счет перитрихиально или полярно расположенных жгутиков, или неподвижные. Облигатные аэробы. Довольно требовательны к субстратам для роста. Почти все виды нуждаются в отдельных витаминах, в первую очередь в пантотеновой кислоте, однако есть формы, способные к синтезу всех факторов роста.
К числу окисляемых соединений относятся одноатомные спирты, содержащие от 2 до 5 углеродных атомов, а также многоатомные спирты — производные сахаров. Окисление первичных спиртов приводит к образованию кислот. Например, этанол с помощью соответствующих дегидрогеназ окисляется до ацетата:
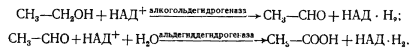
Вторичные спирты окисляются до кетонов:
Многоатомные спирты окисляются этими бактериями в альдозы и кетозы, например: сорбит сорбоза; глицерин диоксиацетон. Альдозы и кетозы могут далее окисляться в соответствующие кислоты. Метаболизирование cахаров осуществляется по окислительному пентозофосфатному пути.
Круг окисляемых соединений различен для разных представителей, входящих в эту группу
С точки зрения характеристики энергетических возможностей уксуснокислых бактерий важно подчеркнуть, что у них развилась удивительная способность воздействовать на определенные химические группировки» осуществляя их одно- или двухступенчатое окисление. Наиболее характерна способность этих бактерий окислять этиловый спирт в уксусную кислоту, давшая название всей группе в целом
Дальнейшая судьба полученных в результате неполного окисления продуктов различна. Некоторые уксуснокислые бактерии не способны к последующим превращениям образовавшихся соединений, и их окислительные способности, следовательно, весьма ограничены. Эти бактерии, объединенные в род Gluconobacter (единственный вид G. oxydans), глюкозу окисляют до глюконовой кислоты, этанол — только до ацетата, который дальше не может ими окисляться из-за отсутствия «замкнутого» ЦТК.
Вторую группу составляют бактерии, способные к полному окислению органических субстратов до CO2 и H2O. В этом случае образовавшаяся уксусная кислота представляет собой лишь промежуточный этап, и после исчерпания из среды исходного субстрата бактерии начинают медленно окислять уксусную кислоту, включая ее в механизм конечного окисления — ЦТК. Бактерии этой группы объединены в род Acetobacter, типичным представителем которого является A. peroxydans.
Электроны от окисляемых субстратов поступают в дыхательную цепь и далее через систему переносчиков передаются на O2, служащий обязательным конечным акцептором электронов. Электронный транспорт приводит к генерированию DmH + .
Место включения электронов в дыхательную цепь определяется ферментом, катализирующим соответствующую окислительную реакцию. Если окисление катализируется НАД-зависимой дегидрогеназой, электроны (водород) передаются на НАД + и с него на переносчики, локализованные на мембране, что открывает возможность для сопряжения электронного транспорта с тремя трансмембранными перемещениями протонов и, соответственно, синтезом 3 молекул АТФ. Недавно у представителей родов Acetobacter и Gluconobacter были обнаружены дегидрогеназы, содержащие в качестве простетической группы соединение из группы хинонов, способные принимать и отдавать 2 атома водорода. Хинонсодержащие дегидрогеназы локализованы на внешней стороне ЦПМ, где и происходит окисление этанола и других соединений. Электроны поступают в дыхательную цепь на уровне цитохромов, а протоны выделяются в периплазматическое пространство.
Уксуснокислые бактерии часто развиваются вслед за дрожжами, используя продукт спиртового брожения как субстрат для роста. Применяются в микробиологической промышленности для получения столового уксуса и в производстве аскорбиновой кислоты (на этапе окисления сорбита в сорбозу).













